简介
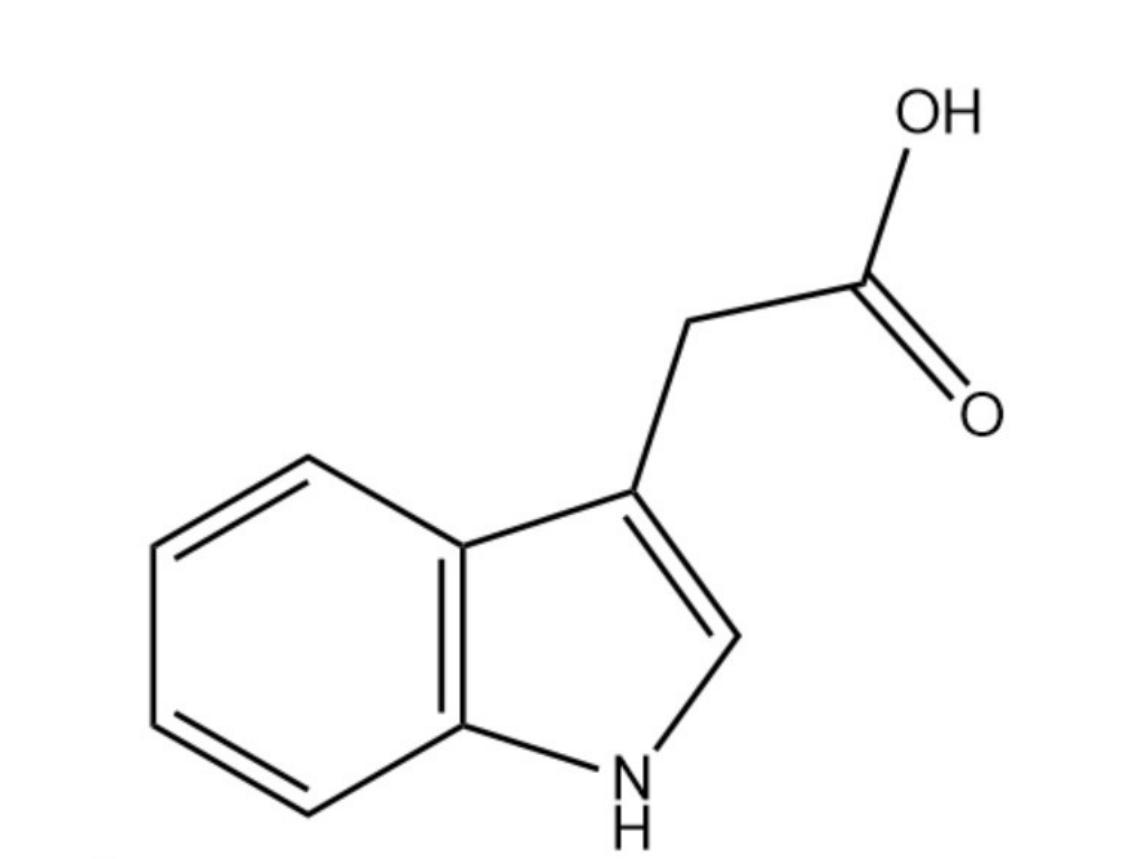
生长素即吲哚乙酸,分子式为C10H9NO2,是最早发现的促进植物生长的激素。英文来源于希腊文auxein(生长)。吲哚乙酸的纯品为白色结晶,难溶于水。易溶于乙醇、乙醚等有机溶剂。在光下易被氧化而变为玫瑰红色,生理活性也降低。植物体内的吲哚乙酸有呈自由状态的,也有呈结合(被束缚)状态的。后者多是酯的或肽的复合物。植物体内自由态吲哚乙酸的含量很低,每千克鲜重约为1-100微克,因存在部位及组织种类而异,生长旺盛的组织或器官如生长点、花粉中的含量较多。
从色氨酸开始,其途径有5个。植物生长素存在于西葫芦中,存在于某些十字花科植物中,存在于番茄中。生长素的降解,最明显的是在光下很容易发生光氧化而被破坏。汤玉玮和J.邦纳于1947年发现植物组织中有些氧化酶能降解吲哚乙酸,称为吲哚乙酸氧化酶。
产生部位
生长素在植物体内分布很广,几乎各部位都有,但不是均匀分布的,在某一时间,某一特定部位的含量是受几方面的因素影响的。大多集中在生长旺盛的部分(胚芽鞘、芽和根尖的分生组织、形成层、受精后的子房、幼嫩种子等),而趋向衰老的组织和器官中则甚少。
运输
极性运输 (Polar Transport)
生长素主要是在植物的顶端分生组织中合成的,然后被运输到植物体的各个部分。生长素在植物体内的运输是单方向的,只能从植物体形态学上端向形态学下端运输,在有单一方向的刺激(单侧光照)时生长素向背光一侧运输,其运输方式为主动运输(需要载体和ATP)。
非极性运输(Non polar transport)
在成熟组织中,生长素可以通过韧皮部进行非极性运输。
合成
IAA的合成前体:色氨酸(tryptophan,Trp)。其侧链经过转氨、脱羧、氧化等反应。锌是色氨酸合成酶的组分,缺锌时导致由吲哚和丝氨酸结合而形成色氨酸的过程受阻,色氨酸含量下降,从而影响IAA的合成。生产上常引起苹果、梨等果树的小叶病。
结合
植物体内生长素有两种形式:游离型:有生物活性,束缚型:活性低。
在体内,吲哚乙酸常常与天门冬氨酸结合成为吲哚乙酰天冬氨酸酯。还可与肌醇结合成吲哚乙醇肌醇。与葡萄糖结合成吲哚乙酰葡萄糖苷。与蛋白质结合成吲哚乙酸—蛋白质络合物。束缚型的生长素可能是生长素在细胞内的一种贮存形式,也是减少过剩生长素的解毒方式,在适当的条件下(pH9-10),它们可转变为游离型,经运输转移到作用部位起作用。
正在生长的种子中生长素的量也多,但完全成熟以后,大部分以束缚态贮藏起来。种子中以束缚态存在,萌发时转变为游离型。
降解
生长素的降解(Degradation of IAA)
①酶氧化降解:吲哚乙酸氧化酶分解
植物体内生长素常处于合成与分解的动态平衡中。吲哚乙酸氧化酶(IAA oxidase)是一种含Fe的血红蛋白。IAA经酶解后形成3—羟基甲基氧吲哚和3—甲基氧吲哚。此反应要在O2存在下,以Mn和一元酚作辅助因子,吲哚乙酸氧化酶才表现活性。
②光氧化分解:
X-光,紫外光,可见光对IAA都有破坏作用,分解产物也是3-亚甲基氧化吲哚和吲哚醛。但机制尚不清楚,在试管里,植物的某些色素,如核黄素,紫黄质等能大量吸收蓝光,并促进IAA的光氧化分解。
植物体内生长素存在的两种形式间的转化或吲哚乙酸氧化酶对IAA的氧化分解都是植物对体内生长素水平的自动调节,对植物生长的调控是有重要意义的。
研究历史
1880年C.R.达尔文及其子在最后出版的著作《植物运动的本领》中阐明,禾本科的加那利草的胚芽鞘被切去顶端就失去向光性响应能力。他的解释是:当幼苗从侧面受光时,顶端产生的影响向下传送,造成向光与背光两侧生长速度不同,从而引起向受光一侧的弯曲,因而切去顶端后就不呈现向光性响应。
1928年F.W.温特用实验证明胚芽鞘尖端有一种促进生长的物质,称之为生长素。它能扩散到琼胶小方块中,将所得小方块放回到切去顶端的胚芽鞘切面的一侧,可以引起胚芽鞘向另一侧弯曲。而且弯曲度大致与所含促进生长的物质的量成正比。这个实验不但证明了促进生长物质的存在,而且创造了著名的测定生长素的“燕麦试法”。
1933年F.克格尔从人尿和酵母中分离出吲哚乙酸,它在燕麦试法中能引起胚芽鞘弯曲以后,证明吲哚乙酸即是生长素,普遍存在于各种植物组织之中。
生理作用
生长素最明显的作用是促进生长,但对茎、芽、根生长的促进作用因浓度而异。三者的最适浓度是茎>芽>根,大约分别为每升10E-5摩尔、10E-8摩尔、10E-10摩尔。植物体内吲哚乙酸的运转方向表现明显的极性,主要是由上而下。植物生长中抑制腋芽生长的顶端优势,与吲哚乙酸的极性运输及分布有密切关系。生长素还有促进愈伤组织形成和诱导生根的作用。
生长素有多方面的生理效应,这与其浓度有关。低浓度时可以促进生长,高浓度时则会抑制生长,甚至使植物死亡,这种抑制作用与其能否诱导乙烯的形成有关。生长素的生理效应表现在两个层次上。
在细胞水平上,生长素可刺激形成层细胞分裂;刺激枝的细胞伸长、抑制根细胞生长;促进木质部、韧皮部细胞分化,促进插条发根、调节愈伤组织的形态建成。
在器官和整株水平上,生长素从幼苗到果实成熟都起作用。生长素控制幼苗中胚轴伸长的可逆性红光抑制;当吲哚乙酸转移至枝条下侧即产生枝条的向地性;当吲哚乙酸转移至枝条的背光侧即产生枝条的向光性;吲哚乙酸造成顶端优势;延缓叶片衰老;施于叶片的生长素抑制脱落,而施于离层近轴端的生长素促进脱落;生长素促进开花,诱导单性果实的发育,延迟果实成熟。
作用机理
激素作用的机理有各种解释,可以归纳为二:
一、是认为激素作用于核酸代谢,可能是在DNA转录水平上。它使某些基因活化,形成一些新的mRNA、新的蛋白质(主要是酶),进而影响细胞内的新陈代谢,引起生长发育的变化。
二、则认为激素作用于细胞膜,即质膜首先受激素的影响,发生一系列膜结构与功能的变化,使许多依附在一定的细胞器或质膜上的酶或酶原发生相应的变化,或者失活或者活化。酶系统的变化使新陈代谢和整个细胞的生长发育也随之发生变化。此外,还有人认为激素对核和质膜都有影响;或认为激素的效应先从质膜再经过细胞质,最后传到核中。
虽然对激素作用机理有不同的解释,但是,无论哪一种解释都认为,激素必须首先与细胞内某种物质特异地结合,才能产生有效的调节作用。这种物质就是激素的受体。
1.激素受体:植物激素受体是指能与植物激素专一地结合的物质。这种物质能和相应的物质结合,识别激素信号,并将信号转化为一系列的生理生化反应,最终表现出不同的生物学效应。受体是激素初始作用发生的位点。所以,了解激素受体的性质及其在细胞内的存在位置,是研究激素作用机理的重要内容之一。激素受体是一种蛋白质,它们可能定位于细胞质膜,也可能定位于细胞核或细胞质。由于植物体内具有多种激素,因此,必然可能有多种激素受体,并存在于细胞的不同部位。
2.生长素最基本的作用是促进细胞的伸长生长,这种促进作用,在一些离体器官如胚芽鞘或黄化茎切段中尤为明显。生长素为什么能促进细胞的伸长生长,又以什么方式起作用的?
植物细胞的最外部是细胞壁,细胞若要伸长生长即增加其体积,细胞壁就必须相应扩大。细胞壁要扩大,就首先需要软化与松弛,使细胞壁可塑性加大,同时合成新的细胞壁物质,并增加原生质。实验证明,用生长素处理燕麦胚芽鞘,可增加细胞壁可塑性,而且在不同浓度的生长素影响下,其可塑性变化和生长的增加幅度很接近,这说明生长素所诱导的生长是通过细胞壁可塑性的增加而实现的。生长素促进细胞壁可塑性增加,并非单纯的物理变化,而是代谢活动的结果。因为,生长素对死细胞的可塑性变化无效;在缺氧或呼吸抑制剂存在的条件下,可以抑制生长素诱导细胞壁可塑性的变化。
相互作用
在植物生长发育的过程中,任何一种生理活动都不是受单一激素的控制,而是各种激素相互作用的结果。也就是说,植物的生长发育过程,是受多种激素的相互作用所控制的。例如,细胞分裂素促进细胞增殖,而生长素则促进增殖的子细胞继续增大。又如,脱落酸强烈的抑制着生长,并使衰老的过程加速,但是这些作用又会被细胞分裂素所解除。再如,生长素的浓度适宜时,促进植物生长,同时开始诱导乙烯的形成。当生长素的浓度超过最适浓度时,就会出现抑制生长的现象。研究激素之间的相互关系,对生产实践有着重要意义。
应用领域
促进生长
生长素(IAA)对营养器官纵向生长有明显的促进作用。如芽、茎、根三种器官,随着浓度升高,器官伸长递增至最大值,此时生长素浓度为最适浓度,超过最适浓度,器官的伸长受到抑制。不同器官的最适浓度不同,茎端最高,芽次之,根最低。由次可知,根对IAA(生长素)最敏感,极低的浓度就可促进根生长,最适浓度为10-10。茎对IAA敏感程度比根低,最适浓度为10-4。芽的敏感程度处于茎与根之间,最适浓度约为10-8。所以能促进主茎生长的浓度往往对侧芽和根生长有抑制作用。
促进分化
生长素与细胞分裂素配合能引起细胞分裂,而且生长素也能单独引起细胞分裂。如早春树木形成层细胞恢复分裂活动是由顶芽产生的生长素下运而引起的。
生长素对器官建成的作用最明显的是表现在促进根原基形成及生长上。苗木插枝在其基部产生不定根,对木本植物来说,主要是由新的次生韧皮部组织分化,但也可由其它组织分化形成,如形成层、维管射线及髓部。吲哚丁酸(IBA)在生长素中促进生根的效果最好,在应用方面发现IBA(吲哚丁酸)与萘乙酸(NAA)比吲哚乙酸(IAA)稳定,效果更好。
维持优势
正在生长的植物茎端对侧芽的生长有抑制作用,这种现象称为顶端优势。棉花用缩节胺控制顶端生长或打顶后,侧芽大量发生。
抑制离区
棉花与果树落花、落果及落叶,是双子叶植物的普遍现象。棉花的蕾铃脱落,与营养物质的供给有关,也与激素水平有关。当蕾铃柄的基部,远轴端生长素含量高,近轴端生长素含量低时,抑制离层内纤维素酶、果胶酶的活性,因而抑制离层细胞的分离,蕾铃不脱落;反之,当近轴端生长素含量高,远轴端生长素含量低时,则使果胶酶和纤维素酶活性提高,促进离层的分离,致使蕾铃脱落。
促进结实
植物开花受精之后,子房中的生长素含量提高,从而促进子房及其周围组织的膨大,加速了果实的发育。如雌蕊未经受精而子房能及时获得IAA,也能诱导某些植物无籽果实的形成。如在授粉前用生长素喷或涂于柱头上,不经授粉最终也能发育成单性果实。如胡椒、西瓜、番茄、茄子、冬青、西葫芦和无花果等
除草剂
除草剂有两种:1. 选择性除草剂,低浓度促进植物生长,高浓度抑制植物生长,对于生长素浓度双子叶植物较单子叶植物更为敏感,因此可作为单子叶植物田中除去双子叶植物的除草剂。2. 非选择性除草剂:将所有植物都杀死,例如草甘膦。
失重影响
根的向地生长和茎的背地生长是要有地球引力诱导的,是由于在地球引力的诱导下导致生长素分布不均匀造成的。在太空失重状态下,由于失去了重力作用,所以茎的生长也就失去了背地性,根也失去了向地生长的特性。但茎生长的顶端优势仍然是存在的,生长素的极性运输不受重力影响。
生长素类似物
类似物
随着对植物激素的研究,人们也在不断的用人工合成的方法制成一些具有植物激素活性的类似物。这些植物激素类似物,一般叫做植物生长调节剂。植物生长调节剂的种类很多,根据功能的不同,可分为植物生长促进剂(如奈乙酸、2,4-D等)、植物生长抑制剂(如三碘苯甲酸、青鲜素等)和植物生长延缓剂(如短壮素、多效唑等)三类。下面举例简要介绍它们的作用和应用情况。
吲哚丁酸:吲哚丁酸简称IBA。纯品为白色或微黄色的晶体,稍有异臭,不溶于水,能够溶于乙醇、丙酮等有机溶剂中。在使用的时候,可以先把它溶解在少量酒精中,然后再加水稀释到所需要的浓度。它主要用于促进植物的插条生根,尤其对生根作用明显。但是,吲哚丁酸诱发出的根细而长,而奈乙酸诱发出的根比较粗壮,因此,生产中常将这两种植物生长调节剂混合使用。
三碘苯甲酸:三碘苯甲酸简称TIBA,纯品为白色粉末,不溶于水,能溶于乙醇、乙醚等有机溶剂中。三碘苯甲酸能够阻碍生长素在植物体内的运输,抑制茎的顶端的生长,促进侧芽的萌发,从而使植株矮化、分枝增多,并且使开花数和结实数增加。三碘苯甲酸已经广泛应用于大豆生产中,用它的溶液喷施大豆植株,可以使植株变矮,分枝增多,结荚率提高,从而提高大豆的产量。
矮壮素:矮壮素简称CCC,化学名称是2-氯乙基三甲基氯化铵。纯品为白色结晶,易溶于水。它的作用与赤霉素相反,能够抑制细胞伸长,但是不抑制细胞分裂,因而能够使植株变矮,茎秆变粗。矮壮素对于防止水稻和小麦倒伏,阻止棉花蕾铃脱落和提高产量,具有明显的效果。由于矮壮素不容易被土壤固定,也不容易被土壤中的微生物分解,所以直接施用到土壤中效果比较好。
多效唑:多效唑简称PP333。多效唑能够抑制赤霉素的生物合成,减缓植物细胞的分裂和伸长,并且抑制茎秆伸长。多效唑广泛应用于果树、花卉、蔬菜和大田作物,效果显著。例如,对番茄幼苗喷施多效唑后,可以使幼苗矮壮,分枝多。中国食品中农药残留标准GB 2763—2005规定了粮谷中多效唑的残留限量标准(MRL)为0.50 mg/kg。